لاکتوز
|
لاکتوز یا قند شیر یک دی ساکارید با فرمول C12H22O11 است و از واحدهای D- گلوکز و D- گالاکتوز که با پیوند بتا 1و4 به یکدیگر متصلند، تشکیل مییابد. جزو قندهای احیا کننده میباشد و به دو فرم ایزومری آلفا و بتا یافت میشود. |
منابع لاکتوز 
بعد از ساکارز ، فراوانترین دی ساکارید طبیعی ، لاکتوز یا قند شیر است. لاکتوز در طبیعت منحصرا در شیر یافت میشود. این قند ، عمدتا در شیر نشخوارکنندگان وجود دارد. به عنوان مثال %5 در شیر گاو. در شیر انسان ، پس از تبخیر تمام مواد فرار ، این ماده نیمی از ماده جامد باقیمانده یعنی %6,5 را تشکیل میدهد. میزان آن در شیر چربیدار یا پُرچربی بسیار کم است، حدود %1.
ساختار لاکتوز
ساختمان لاکتوز بعد از هیدرولیز مشخص شده است که از زیرواحدهای D- گلوکز و D- گالاکتوز تشکیل میشود. این دو جزء سازنده ، بوسیله پیوند بتا- 1و4 به همدیگر متصل میشوند. بنابراین میتوان لاکتوز را D- ? گالاکتوز پیرانوزیل- (1و4)- D- ?- گلوکوپیرانوزید نیز خواند. چون لاکتوز دارای یک کربن آنومری آزاد در زیرواحد گلوکز است، بنابراین در گروه دی ساکاریدهای احیا کننده طبقهبندی میشود. لاکتوز به دو فرم ایزومری آلفا و بتا وجود دارد.
فرم آلفا و بتای لاکتوز
این قند به دو فرم آلفا یا بتا- D- گلوکز پیرانونزید- (4 و1)- D- ? گالاکتوپیرانوزیل وجود دارد. فرم آلفا و بتا خواص بسیار متفاوتی از یکدیگر دارند، از جمله: فرم آلفا دارای چرخش ویژه 89,4+ و نقطه ذوب 202درجه سانتیگراد و فرم بتا دارای چرخش ویژه 35+ و نقطه ذوب 252 درجه سانتیگراد میباشد. هر دو اینها از نظر علمی جالب و در خور توجه میباشند. تبلور آن از آب ، فقط آنومر ? را میدهد.فرم آلفا که با یک مولکول آب ، به صورت بلورین در آمده است، همان لاکتوز متداول و معمول میباشد که میتواند دهیدراته شود (یعنی آب خود را از دست بدهد). فرم بتا (بدون تثبیت آب) ، از محلول غلیظی در حرارت بالاتر از 94 درجه سانتیگراد به شکل بلورین در میآید که این حرارت ، درجه حرارت بحرانی برای فرایند کریستالیزاسیون میباشد. هیدراته شدن فرم بتا تاکنون مشاهده نشده است.
حلالیت لاکتوز
لاکتوز فقط به میزان کمی محلول است، یعنی حدود ده برابر کمتر در مقایسه با ساکارز در درجه حرارت محیط ، محلول میباشد. حلالیت ساکارز در درجه حرارت محیط ، برابر 180 گرم در 100 گرم میباشد و حلالیت لاکتوز نیز به صورت زیر میباشد.
|
| |||
|
|
فرم ? |
فرم ? |
مخلوط |
|
حلالیت (در 100 گرم آب) در دمای 15 درجه سانتیگراد |
7,3g |
50g |
17g |
|
حلالیت (در هر 100گرم آب) در دمای 100 درجه سانتیگراد |
70g |
95g |
- |
تفاوت موجود در میزان حلالیت در درجه حرارت 100 درجه سانتیگراد کمتر است، زیرا میزان محلولیت لاکتوز به مراتب بسیار بیشتر افزایش مییابد.
ظرفیت شیرین کنندگی لاکتوز
لاکتوز دارای قدرت و ظرفیت شیرین کنندگی ناچیزی میباشد. به میزان یک به شش در مقایسه با ساکارز یا قند معمولی است. بنابراین شیر بسیار شیرین نبوده، رژیمهای غذایی دارای شیر ، قابل تحمل هستند یا بعبارت دیگر به آسانی به مصرف سوخت و ساز و جذب میرسند، زیرا ایجاد احساس ناخوشایند نمینمایند.
کریستالیزاسیون لاکتوز
این قند ، دی ساکاریدی است که جداسازی آن با استفاده از کریستالیزاسیون کاملا راحت و آسان میباشد. لاکتوز ، نمونه جالبی از پلی مورفیسم کریستالین میباشد و در عمل کریستالیزاسیون بطئی و آرام متنوعی دیده میشود که در نهایت به شکل بلورهای سخت پیچیده و طویلی ظاهر خواهد شد. مواد معین و مشخصی ، مانع از روند کریستالیزاسیون میگردند. یک نمونه از این مواد ، میتواند ریبوفلاوین (ویتامین Bsub>2) باشد، البته در غلظتی بالاتر از آنچه که این ماده در شیر وجود دارد. هیدرولیز لاکتوز
همانطور که قبلا ذکر شد، لاکتوز در اثر هیدرولیز به دو جزء سازنده اش یعنی گلوکز و گالاکتوز تبدیل میشود. هیدرولیز شیمیایی برای لاکتوز در مقایسه با ساکارز مشکلتر بوده، حرارت دادن آن همراه با یک اسید ، ضروری میباشد و همچنین در هیدرولیز لاکتوز پدیده واژگونی یا اینورسیون (Inversion) که در هیدرولیز ساکارز اتفاق میافتد، وجود ندارد. هیدرولیز آنزیمی مشکلات و مسائلی دارد. آنزیم لاکتاز (بتا- گالاکتوزیداز) به طرز وسیع و گسترده ای یافت نمیشود، هر چند که تعدادی از مخمرهای کمیاب ، این آنزیم را دارا هستند.
توضیح اینکه Klyve remyces Fragilisدر صنعت بکار میرود. این آنزیم ، در قسمت ژژنوم روده ترشح میگردد و بعد از ترشح ، با سرعتهای متفاوتی با توجه به نژادهای متفاوت بشری ناپدید میگردد. در نژادهای رنگینپوست ، زوال ماده مذکور ، زود بوقوع میپیوندد، بعضی وقتها در حدود 3 سال ، در حالیکه در افراد سفیدپوست نسبتی از افراد را مشاهده میکنیم که بیهیچ گونه شکی دارای لاکتاز به میزان بالایی میباشند. بنابراین عارضه ای بنام آلاکتاسیک در میان افراد بالغ که متعلق به نژاد سفید پوست نیستند، رایج و معمول است که مربوط به کمبود لاکتاز و جذب ناصحیح لاکتوز در بدن میباشد.
البته این عارضه در نوزادان متعلق به هر نژادی نادر و کمیاب میباشد. شکل حاد دیگری از عدم توانایی درجذب و متابولیسم مواد غذایی در بچههایی وجود دارد که آنزیم لازم برای تبدیل گالاکتوز به گلوکز ندارند، در اینجا جذب بیشتر منجر به تولید گالاکتوسامیا میگردد و چنین اختلالی در متاولیسم ، نادر بوده و بندرت اتفاق میافتد و باید توجه داشت که گالاکتوز باید به صورت گلوکز تغییر شکل یابد تا بتواند وارد چرخههای بیولوژیکی گردد و همچنین جذب مقادیر فراوانی از گالاکتوز ممکن است که مسلما بیماری آب مروارید باشد.


هسته
|
هسته محل ذخیره اطلاعات ژنتیکی و مرکز کنترل سلول یوکاریوتی است. محتویات هسته در یوکاریوتها توسط غشای هسته احاطه شده است و اندامکی به نام هسته را بوجود آورده است. ولی چون سلولهای پروکاریوتی فاقد غشای هسته هستند بدون هسته محسوب میشوند. |
اطلاعات اولیه 
هسته در سال 1831 توسط Robert Brown در سلولهای اپیدرمی ثعلبیان کشف شد و به عنوان بخشی متراکم ، پایدار و موجود در همه سلولها در نظر گرفته شد. هسته یک ساختار فشرده با پیچ و تابهای زیادی را داراست و با پروتئین همراه میباشد. به چنین مجموعه فشردهای همراه با پروتئین ، کروماتین میگویند. هسته واجد غشای دو لایهای موسوم به پوشش هستهای است و در این پوشش حفرهها یا روزنههایی موسوم به منفذ پیچیده هستهای است که از طریق آنها عمل تبادل مواد بین هسته و سیتوپلاسم انجام میگیرد. برای بررسی ساختمان عمومی هسته میتوان از میکروسکوپ نوری و میکروسکوپ الکترونی استفاده کرد.
|
عکس پیدا نشد |
شکل و محل و تعداد هسته در سلولها
هسته در بیشتر سلولها کروی یا بیضوی است. در سلولهای پارانشیمی بالغ گیاهان عدسی شکل ، در سلولهای عضلانی مخطط جانوران و سلولهای پروکامبیومی گیاهان استوانهای شکل ، در سلولهای آبکش بالغ و سلولهای انگل زده چند بخشی است. در عدهای از سلولها هسته چند بخشی است مثل گویچههای سفید خون چند هستهای و یا سلولهای استئوکلاست (استخوان خوار). در بیشتر سلولها هسته در مرکز قرار دارد. در سلولهای گیاهی به علت رشد واکوئلها ، هسته در کنار غشا قرار میگیرد و در سلولهای عضلانی مخطط هسته در بخشهای کناری قرار دارد.
در جلبک استابولاریا هسته در بخش ریزوئیدی (ریشه نما) یا مجاور با آن قرار دارد. اغلب سلولها دارای یک هسته هستند. با وجود این برخی جانداران ابتدایی و یا سلولهای جانداران پیشرفته بیش از یک هسته دارند. برای مثال حدود 20 درصد از سلولهای کبدی و یا عده زیادی از سلولهای ریسه قارچها دو هستهای هستند. سلولهای عضلانی مخطط ساختمان سنوسیتی دارند یعنی در یک سیتوپلاسم مشترک چندین هسته پراکنده است. این سلولها ابتدا یک هستهای بودهاند که به دلیل تقسیمات مکرر هسته بدون آنکه سیتوپلاسم تقسیم شود به حالت سنوسیتی درآمدهاند.
|
نسبت حجم هسته به حجم سیتوپلاسم را نسبت نوکلئوپلاسمی میگویند. این نسبت برای سلولهایی که در یک مرحله رشد و در شرایط مشابه باشند ثابت است. |
هسته شامل پوشش هستهای ، شیره هسته ، اسکلت هستهای ، کروماتین و بالاخره هستک میباشد.
پوشش هستهای
اطراف هسته سلولهای یوکاریوتی را پوشش هستهای شامل غشای بیرونی ، غشای درونی ، فضای بین دو غشا و منافذ هستهای پوشانیده است.
از دو لایه فسفولیپیدی و پروتئینهای پراکنده در بین آنها تشکیل شده است که شباهت زیادی به غشای شبکه آندوپلاسمی دارد و در سطح آن ریبوزومها قرار گرفتهاند. بخشهایی از غشای بیرونی با شبکه آندوپلاسمی دانهدار ، پیوستگی دارد و منشا غشای هسته نیز از شبکه آندوپلاسمی دانهدار است.
فضای بین دو غشا
فضای بین دو غشای یا فضای دور هستهای فضایی به وسعت 60 تا 100 آنگستروم است که وسعت آن در همه جای پوشش هستهای یکنواخت نیست. در برخی نواحی وسیعتر و در محل منافذ یا سوراخهای هستهای که دو غشا پوشش هستهای بهم میرسند وسعت فضای دور هستهای به صفر میرسد.
غشای داخلی
غشایی زیستی به ضخامت حدود 60 تا 70 آنگستروم ، شبیه غشای شبکه آندوپلاسمی و فاقد ریبوزوم است. غشای داخلی با واسطه پروتئینهای لامینایی با کروماتین ارتباط دارد.
|
عکس پیدا نشد |
منافذ غشای هسته
در پوشش هستهای ساختمانهای پروتئینی فعال و ویژهای به اسم منافذ هستهای وجود دارد. وجود این منافذ بوسیله هرتویگ در سال 1876 برای اولین بار پیشبینی شد. قطر منافذ به اندازهای است که به مولکولهای پروتئین ، انواع RNAها و حتی زیر واحدهای ریبوزومی اجازه عبور میدهد. پروتئینهای سیتوپلاسمی که وارد هسته میشوند از جمله پروتئینهای هیستونی دارای یک بخش نشانه هستند که به کمک آن از بازگشت آنها به سیتوپلاسم جلوگیری میشود. منافذ هستهای عبور یونهای منفی را تسهیل میکنند.
منافذ هستهای ساختمانهای دائمی و پایدار نیستند و متناسب با نیاز سلول ایجاد یا ناپدید میشوند. در سلولهای با فعالیت متابولیکی بالا که مبادله مواد بین هسته و سیتوپلاسم زیاد است تعداد منافذ هسته نیز زیاد است و در سلولهایی که تبادلات هسته و سیتوپلاسم کم است تعداد منافذ کاهش مییابد. هر منفذ بوسیله مجموعهای از ذرات متراکم احاطه شده است. این ساختمانهای پروتئینی را بر روی هم مجموعه منفذی یا منفذ پیچیده هستهای مینامند که شامل بخشهای زیر است.
- یک حلقه یا آنولوس که از 8 پروتئین گرانولی کناری تشکیل شده است و در سطح سیتوزولی قرار دارد.
- یک حلقه یا آنولوس که این هم از 8 پروتئین گرانولی کناری تشکیل شده است و در سطح نوکلئوپلاسمی قرار دارد.
- کانال مرکزی یا درپوش که این کانال محل عبور مواد میباشد و در مرکز منفذ قرار دارد.
- منفذ از ترکیباتی به نام Annular Material پر شده و به سمت نوکلئوپلاسم (شیره هسته) و سیتوپلاسم بیرون زده است.
شیره هسته ( نوکلئوپلاسم یا کاریولنف )
شیره هسته مایعی است که درون هسته را پر کرده است و از نظر کلی شبیه سیتوزول و کمی متراکمتر از آن با PH اسیدی است. شیره هسته یا ماتریکس هسته حاوی آب 10 درصد از کل پروتئینهای هستهای ، 30 درصد از کل RNA و 2 تا 5 درصد از کل فسفولیپیدهای هستهای را شامل میشود. مقدار کمی لیپید و نیز مقداری گلوسیدهای (قندها) موثر در تشکیل نوکلئوتید مثل ریبوز و دزوکسی ریبوز در آن وجود دارند.


در شیره هسته یونهای
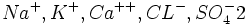{kind=link}
اسیدهای چرب
|
اسیدهای چرب به صورت آزاد به مقدار ناچیزی در سلولها و بافتها دیده میشوند. این ترکیبات که غنی از اکسیژن و کربن و هیدروژن هستند قسمتی از واحد ساختمانی بیشتر لیپیدها مانند چربیهای خنثی ، فسفوگلیسریدها ، گلیکولیپیدها و استرهای کلسترول را تشکیل میدهند. |
دیدکلی 
چربیها یا لیپیدها ترکیبات آلی غیر محلول در آب هستند که میتوان آنها را بوسیله حلالهای غیر قطبی نظیر کلروفرم ، اتر و بنزن از سلولها استخراج کرد. لیپیدها استرهای یک یا چند اسید با الکلها میباشند. بعضی از لیپیدها در ساختمان دیواره و غشای سلولی شرکت داشته و برخی دیگر ماده ذخیرهای انرژیزا را در داخل سلول تشکیل میدهند. اسیدهای چرب به صورت آزاد به مقدار ناچیزی در بافتها دیده میشوند. ساختمان اسیدهای چرب
تاکنون بیش از هفتاد اسید چرب از نسوج گوناگون جدا کردهاند که همگی دارای زنجیره هیدروکربنی طولانی با یک عامل کربوکسیل انتهایی هستند. بعضی از آنها اشباع شده و برخی دارای یک ، دو یا سه پیوند دوگانه هستند و گروهی در زنجیر خود دارای شاخههای جانبی نیز میباشند. تعداد کربنها در اسیدهای چرب به استثنای چند مورد ، همیشه زوج است و احتمالا علت زوج بودن آسان تر بودن سنتز آنها در نزد جانداران است. فرمول کلی اسیدهای چرب CH3-(CH2)n-COOH است که n از صفر تا 30 الی 40 تغییر میکند.
خواص اسیدهای چرب
اسیدهای چرب غیر اشباع دارای نقطه ذوب پایینتری نسبت به اسیدهای چرب اشباع هستند و هر چه تعداد کربن اسید چرب بیشتر شود، نقطه ذوب بالاتر میرود. پیوند دوگانه در اسیدهای غیر اشباع بیشتر بین کربن 9 و 10 بوده و این پیوند دوگانه تقریبا در تمامی اسیدهای چرب غیر اشباع طبیعی در وضعیت ایزومر هندسی سیس (Cis) است. اسیدهای چرب با زنجیر طویل در آب غیر محلول هستند. ولی در قلیایی محلولاند و تشکیل صابون سدیم یا صابون پتاسیم میدهند. اسیدهای چرب غیر اشباع به سهولت اکسید میشوند. تند شدن چربیها بر اثر اکسید شدن و ایجاد عوامل اسیدی و آلدئیدی در چربیها است.
ساختمان و خواص انواع چربیها
چربیهای خنثی (آسیل گلیسرولها)
ترکیب اسید چرب و گلیسرول را آسیل گلیسرول یا گلیسرید مینامند. گلیسرول الکلی است که بیش از سایر الکلها در ساختمان لیپیدها دیده میشود و دارای 2 عامل الکلی نوع اول و یک عامل الکلی نوع دوم است. برحسب آن که یک ، دو یا سه اسید چرب با عوامل الکلی گلیسرول ترکیب شده باشد، به ترتیب مونو ، دی و تری آسیل گلیسرول بدست میآید. نقطه ذوب چربیها خنثی بستگی به نقطه ذوب اسید چرب ترکیبی آنها دارد. تری آسیلها در آب نسبتا نامحلول بوده و تشکیل میسلهای کامل نمیدهند. در حالی که دی آسیل گلیسرولها که تا حدودی قابل یونیزه شدن هستند، به سهولت تشکیل میسل میدهند.
فسفو گلیسریدها
این لیپیدها که به نام گلیسرول فسفاتید نیز موسومند، بیشتر در غشاهای سلولی وجود دارند و فقط به مقدار خیلی جزئی در چربیهای ذخیرهای یافت میشوند. الکل این لیپیدها گلیسرول است که یکی از عوامل الکلی نوع اول آن توسط اسید فسفریک استریفیه شده است. تمام فسفر گلیسریدها دارای یک انتهای قطبی و دو انتهای طویل غیر قطبی میباشند و به این جهت آنها را لیپیدهای قطبی یا آمفی پاتیک گویند.
پلاسمالوژنها
یک گروه فرعی از فسفر گلیسریدها هستند که در آنها به جای یک مولکول اسید چرب یک آلدئید چرب قرار گرفته است. این ترکیبات در غشای سلولهای عضلانی و عصبی فراوانند.
اسفنگو لیپیدها
اینها در غشای سلولهای گیاهی و سلولهای حیوانی و در بافتهای عصبی و مغز به مقدار فراوان وجود دارند. این ترکیبات در اثر هیدرولیز ایجاد یک مولکول اسید چرب و یک مولکول الکل آمینه غیر اشباع به نام اسفنگوزین میکنند. در اسفنگو لیپیدها گلیسرول وجود ندارد. ترکیب اسفنگوزین و اسید چرب را سرامید گویند. اسفنگو میلین ترکیب سرامید با فسفو کلین میباشد که فراوانترین اسفنگو لیپیدها است.
گلیکو لیپیدها
این ترکیبات دارای یک انتهای قطبی هیدروفیل (آبدوست) قندی میباشند. برخی شامل اسفنگوزین و برخی دیگر شامل گلیسرول هستند. قندهای آن شامل D- گلوکز و D- گالاکتوز است. گانگلیوزیدها ترکیبات دیگری از دسته گلیکو لیپیدها هستند. این ترکیبات عبارتند از گلیکو اسفنگو لیپیدهایی که دارای انتهای قطبی متشکل از قندهای مرکب نظیر اوزامین و اسید سیالیک میباشند. این ترکیبات در غشای سلولی بویژه در سلولهای عصبی زیاد دیده میشوندسربروزیدها
این ترکیبات را میتوان در گروه گلیکو لیپیدها و یا اسفنگو لیپیدها طبقه بندی کرد. زیرا دارای قند و اسفنگوزین هستند. این دسته از لیپید بیشتر در غشای سلولهای عصبی بویژه در غلاف میلین و همچنین در گویچههای قرمز خون ، گویچههای سفید خون و اسپرم دیده میشوند.
مومها
مومها از نظر ساختمانی و خواص ، شبیه آسیل گلیسرولها هستند. ولی الکلهای آنها دارای زنجیره کربنی طویل هستند که تنها شامل یک عامل الکل میباشند. این ترکیبات در غشای محافظ پوست ، پر و جدار برگها و بعضی میوهها و پوسته خارجی بیشتر حشرات دیده میشوند.
لیپیدهایی که صابونی نمیشوند
لیپیدهایی که تاکنون مورد بحث قرار گرفتند قابل صابونی شدن بوده یعنی با قلیاییها و در اثر حرارت ایجاد صابون میکنند. در سلولها مقدار کمتری از نوع دیگر چربیها موجودند که غیر قابل صابون شدن هستند. دو گروه اصلی از این لیپیدها یکی استروئیدها و دیگری ترپنها هستند.
- استروئیدها: ساختار استروئیدها از سه حلقه شش ضلعی فنانترن و یک حلقه پنج ضلعی هستند. از میان مهمترین استروئیدهایی که در طبیعت دیده میشوند، اسیدهای صفراوی ، هورمونهای جنسی ، هورومونهای قسمت قشری غدد فوق کلیوی و ویتامین D و کلسترول را میتوان نام برد. استروئیدها به مقدار بسیار کم در سلولها موجودند و فقط یک نوع آنها که بطور کلی استرول نامیده میشود، بسیار فراوان است. کلسترول فراوانترین نوع استرولها در بافتهای حیوانی است. کلسترول در گیاهان دیده نمیشود. استرول موجود در گیاهان به نام فیتوسترولها مشهورند. قارچها و مخمرها حاوی استرولهایی به نام میکو استرول هستند. در این دسته ارگوسترول را میتوان نام برد که به ویتامین D تبدیل میشود.
- ترپنها: به مقدار جزیی در سلولها وجود دارند. ترپن ممکن است دارای ساختمان خطی یا حلقوی باشد. ویتامین A ، کاروتنها ، بیشتر اسانسها و ویتامین E و ویتامین K از ترپنها مشتق میشوند.
لیپو پروتئینها
لیپیدهای قطبی با پروتئینهای خاصی ترکیب شده و لیپید پروتئینها را میسازند. مانند لیپو پروتئینهای پلاسمای خون که نقش حامل مواد مختلف را دارا میباشند. برخی لیپیو پروتئینها دارای لیپیدهای خنثی مانند گلیسرول و آسترهای آن هستند. این ترکیبات وسیله انتقال چربیها از روده کوچک به کبد به بافت چربی و دیگر بافتها هستند. طبقه بندی لیپو پروتئینها بر مبنای وزن مخصوص آنها انجام میگیرد.
پروستا گلاندینها
این ترکیبات که برای نخستین بار در مایع منی و غده پروستات یافت شدند، از یک اسید چرب غیر اشباعی 20 کربنه به نام اسید آراشیدونیک مشتق شدهاند. این ترکیبات انواع مختلفی دارند که تاکنون متجاوز از 14 نوع پروستاگلاندین در مایع منی انسان و تعداد دیگری در سایر بافتها یافت شده است. پروستاگلاندینهای گروه E ، گروه F و گروه A و B نمونه این ترکیبات هستند. پروستاگلاندینها دارای اثرات فیزیولوژیک مختلفی مانند کاهش فشار خون ، تنظیم عبور یونهای مختلف از غشای سیناپسهای عصبی و خنثی سازی اثر برخی هورمونها میباشند.
میسلها
لیپیدهای قطبی مانند اسیدهای چرب ساده و صابونها در محلولهای آبگون پخش شده و میسلها را بوجود میآورند که در آنها انتهای هیدروکربنی چون آبگریزند (هیدروفوب) از محلول آبگون رانده شده و ایجاد یک فاز آبگریز داخلی میکند و در نتیجه انتهای آب دوست (هیدروفیل) آنها به طرف خارج گرایش مییابد. تری آسیل گلیسرولها و کلسترول خود به خود تشکیل میسل نداده ولی در ساختمان مسیلی وارد شده و میسلهای مخلوط را میسازند. میسلها ممکن است تک لایه یا دو لایه باشند.